- КРОВЕТВОРЕНИЕ
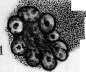
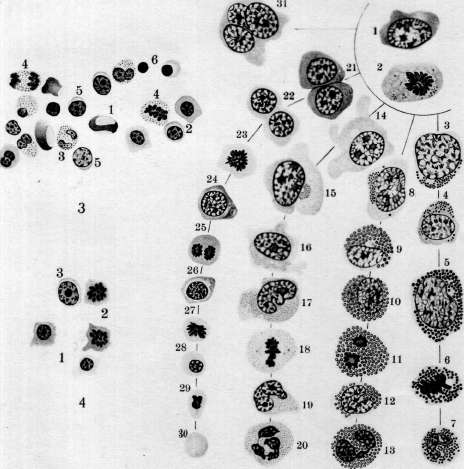
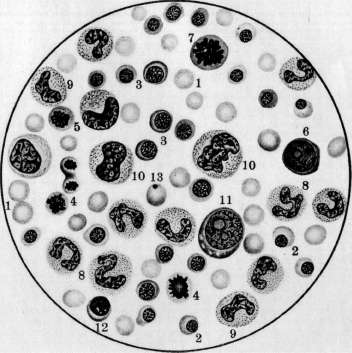
- КРОВЕТВОРЕНИЕ, КРОВЕТВОРНЫЕ ОРГАНЫ. Под кроветворением, или гемо-п о э з о м (от греч. haima—кровь и poesis— творение), понимают процессы образования форменных элементов крови. К. происходит в т. н. кроветворных органах и состоит в развитии, размножении и созревании кро-' вяных телец. Оно протекает непрерывно, т. к. форменные элементы крови после кратковременного функционирования подвергаются изнашиванию и распаду. При ди-ференцированном К. можно отделить процессы развития красных кровяных шариков (эритропоэз) от развития белых кровяных шариков зернистых (гранулопоэз) и незернистых (лимфопоэз). Эмбриональное кроветворение. Кровяные клетки эмбриона являются производными мезенхимы, одновременно с развитием к-рой начинается К. Первым кроветворным органом являются кровеносные сосуды в сосудистом поле (см. Area vasculosa), следовательно вне тела зародыша—период внеэмбриональ-ного К.; затем главным кроветворным органом делается печень—период печоночного К., или домозговой период, и наконец несколько позднее начинает функционировать костный мозг—костномозговой период. Кроме того в К. зародыша принимают участие мезенхима различных органов, селезенка и лимфоидная ткань.—В неэмбриональное К. начинается в самый ранний период развития зародыша, одновременно с развитием мезенхимы: в area ораса желточного мешка у млекопитающих, птиц, рептилий и селяхий образуются отдельные скопления клеток мезенхимы, т.н. кровяные островки Вольфа, центральные клетки к-рых закругляются, а наружные уплощаются, образуя эндотелиальную оболочку. Число и объем кровяных островков быстро увеличивается; они окружают со всех сторон развивающийся зародыш, располагаясь в т. н. сосудистом поле (area vasculosa). Клеточные тяжи, которыми кровяные островки соединяются друг с другом, превращаются в тонкостенные трубки, благодаря чему возникают первичные кровеносные сосуды с плавающими в жидкости круглыми клетками — первичными кровяными клетками (Максимов). У эмбрионов человека по ис- следованиям Шридде (Schridde) сперва появляются пустые зачатки кровеносных сосудов, из стенки к-рых уже вторично происходят свободные плавающие в жидкости клетки. Первичны*е кровяные клетки остаются без изменения самое короткое время, а затем диференцируются в крупные гемоглобин-содержащие клетки — первичные эритро-бласты и далее в эритроциты. Впрочем часть их, по Максимову, сохраняется в недиферен-цированном состоянии, функционируя первые бесцветные кровяные тельца зародыша. Первичные эритробласты и эритроциты служат переносчиками кислорода в ранний период эмбрионального развития и затем постепенно вымирают (у человеческого зародыша в конце 3-го месяца).—Вторым кроветворным органом является печень, кроветворение в которой начинается у человека ; в конце 2-го месяца эмбриональной жизни. Согласно исследованиям Заксера, Молье, Неймана (Saxer, Mollier, Neumann), Максимова из периваскулярной мезенхимы эмбриональной: печени образуются гемоци-тобласты, которые экстраваскулярно дают происхождение вторичным эритробластам и мегакариоцитам. Вторичные эритробласты вполне соответствуют эритробластам взрослого организма и, созревая, дают происхождение вторичным эритроцитам. Грануло-циты в печени также развиваются экстраваскулярно, чаще из мелких форм блуждающих клеток без типичного стадия гемоцито-бласта и миелоцита. Во время среднего периода-беременности кровеобразование в печени достигает наибольшей степени, и ббль-шая часть массы органа состоит из кро-веродной ткани; у новорожденного печень содержит еще очаги эмбрионального кроветворения. Костный мозг как кроветворный орган начинает функционировать позднее печени. Мезенхима тела уже с самых ранних стадиев развития эмбриона служит местом К. Фиксированные мезенхимные клетки и эндотелий кровеносных сосудов округляются, изолируются и дают начало двоякого рода блуждающим клеткам: типа гемоцито-бластов и гистиоцитам (Максимов). Те и другие способны давать в процессе дальнейшего развития одинаковые клетки и могут вероятно переходить друг в друга. Заксер назвал их первичными блуждающими клетками. Из них развиваются разбросанные в мезенхиме гемопоэтические очаги, содержащие эритробласты, гранулоциты и ме-гакариоциты. Первые лимф.узлы появляются в стенке шейного лимф, мешка у человеческого зародыша в 30 мм длиной; позднее они развиваются в других лимф, мешках, а также по ходу лимф, сосудов. Из мезенхимы стенки лимф, мешка происходит образование полиморфных первичных блуждающих клеток, среди к-рых развиваются с самого начала в больших количествах малые лимфоциты, частью путем непосредственного закругления мелких мезенхим-ных клеток, частью при делении более крупных блуждающих клеток. Утверждение Шридде, Тюрка и Негели (Turk, Naegeli) о развитии лимфоцитов из эндотелия лимф. сосудов не подтверждается Максимовым. Кроме того в зачатках лимфатич. узлов из первичных блуждающих клеток могут развиваться гранулоциты, а в более поздних стадиях—отдельные мегакариоциты и мелкие островки из эритробластов. Настоящие фоликулы с зародышевыми центрами развиваются только в постэмбриональной жизни.—Эмбриональная селезенка является местом значительного эритропоэза, продолжающегося до конца эмбриональной жизни, тогда как гранулопоэз отсутствует. И здесь клетки мезенхимы дают происхождение кровяным элементам. У других позвоночных эмбриональное К. протекает в общем по тому же плану, как и у млекопитающих. У птиц в сосудах area vasculosa первичные кровяные клетки в большей своей части диференцируются в 'первичные эритробласты (Данчакова), а часть клеток остается недиференцированной и в дальнейшем функционирует как гемоцитобла-сты. Из них в конце 4-го дня насиживания начинают развиваться вторичные эритробласты, созревающие в ядросодержащие эритроциты. В отличие от млекопитающих у птиц в сосудистом поле развиваются экстраваскулярно из клеток мезенхимы очень многочисленные гранулоциты через стадий гемоцитобласта (см.).. Печень не играет существенной роли в К. В эмбриональном костном мозгу эритропоэз протекает интра-васкулярно, а гранулопоэз—экстраваскулярно, что сохраняется в течение всей жизни. Вместо кровяных пластинок млекопитающих у всех остальных позвоночных в крови циркулируют мелкие клетки—тромбоциты, которые развиваются у птиц в-сосудах костного мозга из малых лимфоцитов через стадий тромбобласта (мегакариоциты здесь отсутствуют). Тогда как эмбриональное К. у рептилий стоит очень близко к птицам, у амфибий она представляет нек-рые особенности: у зародышей лягушки и аксолотля циркулирующая кровь вначале содержит только крупные, богатые желтком и пигментом клетки, большая часть которых превращается в первичные эритробласты. У бесхвостых амфибий (лягушка) важным кроветворным органом в эмбриональном периоде является мезенхима в области предпочки, первичной почки и во влагалище брыжеечной артерии, где происходит лимфо- и гранулопоэз. Во всех кровеносных сосудах эмбриона из малых лимфоцитов развиваются тромбоциты, а из больших—вторичные эритробласты; несколько позднее эритропоэз сосредоточивается в сосудах печени и наконец окончательно устанавливается на всю жизнь в просвете сосудов костного мозга, тогда как лимфо- и гранулопоэз протекают экстра-. васкулярно. У хвостатых амфибий (аксолотль) лимфо- и гранулопоэз происходят в печени, а эритропоэз—в селезенке. Первичные почки здесь не играют роли. Из рыб эмбриональное кроветворение более подробно изучено у селяхий (Максимов), у к-рых оно представляет большое сходства с кроветворением птиц и рептилий. Помимо К. в стенке желточного мешка, в определенных местах мезенхимы тела, а именно—в стенке пищевода (Лейдиговский орган) ив тесном соприкосновении с канальцами, первичной почки и половых желез развиваются очаги лимфо- и гранулопоэза, функционирующие в течение всей жизни. Селезенка является окончательным эритро- и тромбо-цитопоэтическим органом, а в более поздних стадиях эмбрионального развития в ней вокруг артерий развиваются большие скопления лимфоцитов. Кроветворные органы. К. в постэмбриональном периоде происходит в миелоидной ткани, лимфоидной ткани и рет.-энд. аппарате. Миелоидная ткань костного мозга является при нормальных условиях единственным местом развития зернистых лейкоцитов, эритроцитов и кровяных пластинок (см. Костный мозг). В петлях ретикулярной ткани костного мозга лежат свободно клеточные элементы, образующие паренхиму (рис. 1). К ним относятся эри-тробласты, эритроциты, зернистые лейкоциты и их молодые формы, гемоцитобласты и мегакариоциты. Имеются ли в паренхиме малые лимфоциты и моноциты — спорно. В 1 мм3 костного мозга, выжатого .из ребра взрослой собаки, содержится около 500.000 эритробластов и 1.200.000 лейкоцитов (Ти-мофеевский). Лимфаденоидная ткань является местом образования лимфоцитов. Из нее состоят лимф, узлы, белая пульпа селезенки и лимф, фоликулы слизистых оболочек. Соединительнотканный остов ее состоит из ретикулярной ткани, в петлях к-рой лежат лимфоциты (рис. 7).— Рет.-энд. система организма многими авторами рассматривается как самостоятельный кроветворный орган. К ней относятся блуждающие клетки соединительной ткани, находящиеся в покое, ретикулярные клетки костного мозга, селезенки, лимф. узлов, включая сюда и эндотелий, покрывающий лимф, синусы и венозные синусы костного мозга и селезенки, далее Купферовские клетки печени, часть эндотелия венозных ка-пиляров надпочечника и гипофиза. Участие ретикуло-эндотелия в К. не может считаться вполне выясненным. Согласно исследованиям Феррата, Маршана, Герцога (Ferrata, Marchand, Herzog), Хлопина, Сысоева и других при известных условиях из него могут развиваться элементы миелоидной ткани; другие (Максимов) отрицают способность его к гемопоэзу. Исследования Ашо-фа, Кийоно (Aschoff, Kiyono) и Чашина показали, что при достаточно сильной прижизненной окраске животных литиевым кармином отдельные мобилизованные рет.-энд. клетки или гистиоциты, прижизненно окрашенные, появляются в крови, особенно— правого сердца. В связи с этими данными и в виду близкого сходства кровяных гистиоцитов с моноцитами возникло учение о происхождении моноцитов крови из клеток ретикуло-эндотелиальн. аппарата (Aschoff, Kiyono, Schilling). Кроветворение в разные периоды жизни человека представляет некоторые особенности. Интенсивность гемопоэза, свойственная детскому возрасту, сменяется постепенно более медленным темпом регенерации крови у взрослых и значительным ослаблением функций кроветворных орга- нов в старческом организме. У новорожденного кроветворные органы носят еще на себе некоторые черты эмбрионального периода: костный мозг богат гемоцитобласта-ми, в печени имеются остатки эмбрионального гемопоэза, лимфатич. узлы содержат много больших лимфоцитов', лишены еще центров размножения Флемминга. Лимф. узлы в раннем детском возрасте относительно большие, бедны фоликулами, центры размножения в которых начинают обозначаться к 2 месяцам, но отчетливо выступают только к двухлетнему возрасту. Повышенная функция кроветворных органов в детском возрасте отражается на составе периферической крови: в ней мы находим молодые формы красных и белых кровяных телец — эритробласты, полихроматофильные эритроциты, большие лимфоциты. Этот же период жизни характеризуется частым и обильным развитиемэкстрамедулярного кроветворения, легкостью нарушения функций кроветворных органов и более частым мега-лобластическим превращением костного мозга под влиянием различных вредностей, чем у взрослых. В стареющем организме интенсивность гемопоэза падает, красный костный мозг беднеет паренхиматозными элементами, взамен к-рых развиваются жировые клетки; область распространения его уменьшается; особенно ослабляется эритро-поэз; в лимфатических узлах наступают ат-рофич. изменения, часть лимф, узлов подвергается полному запустению и фиброзу. Способность кроветворных органов старика отвечать повышением своей функции на усиленный распад элементов крови в значительной степени ослабляется. Эритропоэз (нормальный и патологический). В норме эритробласты развиваются в костном мозгу преимущественно гомопластическим способом, т. е. путем размножения уже существующих там эритробластов. При своем.развитии в эритроцита эритробласт проходит несколько стадиев, а именно—проэритробласта, поли-хроматофильного эритробласта, нормоблас-та и наконец нормоцита (рисунок 5). При этом базофилия протоплазмы, резко выраженная у проэритробластов, постепенно ослабевает, в протоплазме накопляется НЪ, ядро уменьшается в размерах, теряет ядрышки, получает более грубую структуру. Наконец ядро сморщивается, делается пик-нотичным и затем, по одним авторам, выталкивается из нормобласта, а по другим—подвергается внутриклеточному растворению. Процессы деления путем кариокинеза играют важную роль при таком постепенном созревании эритробласта. При этом размер клеток следующих генераций делается все мельче и мельче, приближаясь к размеру эритроцита (рис. 5). Убыль эритробластов, зависящая от созревания их в эритроциты, пополняется размножением главн. обр. уже гемоглобинсодержащих клеток, тогда как проэритробласты находятся как бы в резерве и только при повышенном эритропоэ-зе начинают вырабатывать в своей протоплазме НЬ и усиленно размножаться. Полихроматофильные эритробласты, развивающиеся непосредственно из проэритро- Рисунок 1. Срез из костного мозга кролика. Тромбоцитопоэз: 1—мегакариоцит с мелкозернистой протоплазмой; 2—просвет синусоида костного мозга; 3—бляшки Биццоцеро, образующиеся из протоплазмы мегакариоцита; 4—эндотелий синусоида. (С препарата из неопубликованной работы Часовникова.) Рисунок 2. Схема генетических отношений элементов соединительной ткани крови (стрелки указывают направление развития): 1—недиференцированная мезенхимная клетка; 2—гемо-цитобласт (большой лимфоцит); 3—фиброцит; 4—гистиоцит; 5—базофильный лейкоцит; 6—эозинофильный лейкоцит; 7—нейтрофильный лейкоцит; 8—эритроцит; 9—мегакариоцит; 10—малый лимфоцит; 11—моноцит. (По Максимову.) Рисунок 3. 20-часовая культура гемоцитобластов из крови больного острой миелоидной лейкемией. Гемопоэз in vitro: 1—эритроцит; 2—эритробласты; 3—сегментированные нейтро-филы; 4—фигуры деления нейтрофильных миелоцитов; 5—гемоцитобласт; 6—ядро дегенерировавшей клетки. Рисунок 4. Размножение эритробластов в такой же культуре: 1—эритробласты; 2—фигуры деления полихроматафильных эритробластов; 3—гемоцитобласт. Рисунок 5. Развитие миелоидных элементов человеческого костного мозга из гемоцитобласта: 1—гемоцитобласт; 2—фигура кариокинеза гемоцитобласта; 3 и 4—базофильные промиело-циты; б—базофильный миелоцит; б—фигура кариокинеза базофильного миелоцита; 7—полиморфноядерный базофил; 8—эозинофильный промиелоцит; 9 и 10—эозинофильные миело-циты; 11—фигура деления эозинофильного миелоцита; 12—эозинофильный метамиелоцит; 13—полиморфноядерный эозинофил; 14 и 15—нейтрофильные промиелоциты; 16 и 17—ней-трофильные миелоциты; 18—фигура кариокинеза нейтрофильного миелоцита; 19—нейтрофильный метамиелоцит; 20—полиморфноядерный нейтрофил; 21—проэритробласты, происшедшие из разделившегося гемоцитобласта; 22—26—полихроматофильные эритробласты, постепенно обогащающиеся гемоглобином; 27—фигура деления нормоблаета; 28—зрелый нюрмобласт; 29—нормобласт с пикнотическим ядром; рядом—свободное вытолкнутое ядро; 30—эритроцит; 31—мегакариоцит. (По Максимову.) Рисунок 6. Срез через костный мозг кролика после внутривенного впрыскивания литиева кармина и туши: 1—нормобласты; 2—полихроматофильные эритробласты в состоянии кариокинеза; 3—проэритробласты; 4—эозинофильные миелоциты; 5—специальные миелоциты; 6—фигура деления специального миелоцита; 7—гемоцитобласт; 8—мегакариоцит; 9—ретикуло-эндотелиальные клетки с протоплазмой, набитой глыбками туши и зернышками литиева кармина; 10—просвет синусоидов; 11—жировая клетка; 12—артерия; 13—эритроциты в просвете синусоида. (По Максимову.) Рисунок 7. Срез из лимфатического узла кролика. Часть фоликула с центром размножения в активном стадии: 1—большие лимфоциты; 2—средние лимфоциты; 3—малые лимфоциты; 4—фигуры деления средних лимфоцитов; 5—индиферентная клетка мезенхимы; 6—макрофаг. Рисунок 8. Мазок костного мозга из ребра малокровной собаки. Резко выраженный эритропоэз: 1—эритроциты; 2—нормобласты; 3—полихроматофильные эритробласты; 4—фигуры деления нормобластов; 5—фигура деления полихроматофильного эритробласта; 6—проэритро-бласт; 7—фигура деления проэритробласта; 8—палочкоядерные нейтрофилы; 9—юные ней-трофилы; 10—нейтрофильные миелоциты; 11—гемоцитобласт; 12—лимфоцит; 13—эритроцит с тельцем Жолли. (К иллюстр. ст. Кроветворение.)
(В
i*^ ^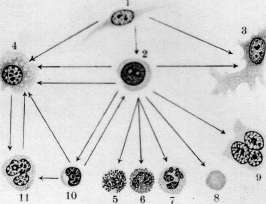
К ст. Кроветворение.

M*>i£**.fb.
ф| # ©р Чадит $ фМк
К ст. Кроветворение.
бластов, имеют более крупные размеры, чем эритроциты, а их протоплазма окрашивается смесью кислых и основных красок в промежуточный тон. Благодаря их крупным размерам их называют иногда макро-бластами. Эритробласты лежат в паренхиме костного мозга небольшими кучками клеток, находящихся в одинаковых стадиях созревания (рис. 6). Механизм проникновения эритроцитов из паренхимы костного мозга в просвет синусов не вполне выяснен.—Г етеропластический зрит* ропоэз, т. е. развитие проэритробластов из индиферентной.клетки, еще не вполне ясен. По взгляду унитаристов (Максимов), гемоцитобласт костного мозга может дифе-ренцироваться в различных направлениях, в том числе и в проэритробласта (рис. 5). При этом кариокинетическое деление является необходимым, т. к. во время него происходит внутренняя перестройка ядра клетки, к-рая становится способной развиваться лишь в эритроцита. По учению дуалистов (Негели) молодой эритробласт (пронормоб-ласт) развивается из индиферентной мезен-химной клетки, а Шридде производит его из эндотелия кровеносных сосудов. Эллер-ман (Ellermann) говорит об особи материнской клетки эритробластическ. ряда, о т. н. эритрогонии. Опыты эксплянтации лейке-мической крови доказывают, что гемоцитобласт способен в некоторых случаях in vitro диференцироваться в сторону эритро-бластов (Тимофеевский и Беневоленская).— Пат. увеличение эритропоэза наблюдается гл. обр. при анемиях (рис. 8). При этом содержание эритробластов в костном мозгу увеличивается, количество митозов возрастает, гетеропластический эритропоэз усиливается, жировой костный мозг замещается красным.. При некоторых особенно тяжелых анемиях, напр. злокачественном малокровии, развивается экстрамедуляр-ное К. (см. ниже гетеротопное кроветворение). При сильных интоксикациях и инфекциях может произойти угнетение эритропоэза: число эритробластов падает, фигуры деления их исчезают (арегенеративные— апластические анемии). Экспериментально подавление эритропоэза удается вызвать повторными кровопусканиями. Уменьшение эритропоэза наблюдается далее при лейкозах, особенно—острых. При злокачественном малокровии наблюдается развитие в костном мозгу наряду с нормобластами очень крупных гемоглобинсодержащих клеток—-мегалобластов и мегалоцитов. Эрлих(ЕпгИсп) высказал взгляд, что мегалобластическое превращение костного мозга является возвратом к эмбриональному К., а мегалобла-сты идентичны с первичными эритробласта-ми. Эта гипотеза нашла поддержку среди многих гематологов. Гетеропластическое развитие мегалобластов не может считаться вполне выясненным. По Негели, они развиваются из мезенхимных клеток, по Фер-рата (Ferrata) — из гемогистиобластов (гистиоцитов), по Ламбену (Lambin)—из ре-тикуло-эндотелия. Кроме того нарушение нормального эритропоэза может выражаться: 1) в распаде ядра эритробластов на отдельные сегменты; 2) в сохранении в эритро- бласте остатков ядерной субстанции (тельца Жолли, кольца Кабота); 3) в полихромазии и базофильной "точечности эритроцитов и наличии в них ретикуло-филаментозной субстанции, окрашиваемой основными красками в нефиксированных препаратах; 4) в недостаточной выработке НЬ. Некоторые из этих отклонений встречаются иногда в нормальной крови, но особенно сильно выражены при анемиях. Лейкопоэз (нормальный и патологический). 3 е р ни сты е лейкоциты развиваются в норме исключительно в костном мозгу преимущественно гомопластическим способом, т. е. путем размножения и созревания молодых зернистых клеток, так наз. миелоцитов (рис. 5, 6 и 8). Эти крупные клетки имеют круглое или бухтообразное ядро и зернистую протоплазму. Соответственно характеру зернистости различают нейтрофильные, эозинофильные и базофиль-ные миелоциты. Созревание миелоцитов в соответствующие сегментированные лейкоциты проходит через несколько стадиев, более подробно изученных для нейтрофи-лов. Ядро клетки делается сперва подковообразным (метамиелоцит Паппенгейма или юный нейтрофил, по Шиллингу), затем получает палочковидную форму в виде изогнутого, густо окрашивающегося жгута (палочкоядерный нейтрофил, по Шиллингу) и наконец распадается на несколько (2—5 и больше) сегментов, соединенных узкими перемычками (сегментированный нейтрофил). Начиная со стадия метамиелоцита, размножение клеток прекращается. Гетеропластический гранулопоэз, являющийся обычным при эмбриональном К., во взрослом организме в норме встречается редко. Считается доказанным, что все 3 вида миелоцитов происходят из общей родоначаль-ной клетки—гемоцитобласта (миелобласта). Другие, менее употребительные названия для этой клетки: лимфоидоцит(Паппенгейм), базофильный миелоцит (Dominici), лимфо-идная костномозговая клетка (Тюрк). Унитаристы (Максимов) помимо названия «гемоцитобласт» обозначают ее также большим лимфоцитом, тогда как дуалисты (Негели) придерживаются названия «миелобласт». Гемоцитобласт—клетка крупных размеров с круглым или слегка бухтообразным ядром, хроматин которого на сухих окрашенных мазках имеет вид тонкой равномерной сеточки с мелкими узловыми утолщениями. В ядре имеется несколько ядрышек. Протоплазма базофильна, незерниста, количество ее невелико. Гемоцитобласт дает положительную реакцию на протеолити-ческий фермент, оксидазу и пероксидазу— в этом видят отличие его от морфологически близко стоящего к нему большого лимфоцита или лимфоблаота, не дающего этих реакций. Во взрослом организме количество гемоцитобластов невелико, у новорожденных их значительно больше, а в эмбриональном периоде они преобладают над остальными клетками. Гемоцитобласт при созревании в миелоцита должен, по Максимову, подвергнуться митозу, во время которого и происходит диференцировка дочерних клеток в том или ином направлении (рис. 5). При этом базофилия протоплазмы уменьшается, количество последней увеличивается, в ней иногда развивается густая азурофильная зернистость, хроматиновые глыбки ядра делаются более грубыми, а ядрышки менее отчетливыми. Затем начинает вырабатываться тот или другой вид специфической зернистости, сперва в незначительном количестве, обычно около ядра на месте бухтообразного углубления. Такие переходные формы от гемоцитобластов к миелоцитам выделяются под названием про-миелоцитов. При дальнейшем созревании промиелоцита азурофильная зернистость, если она имелась, исчезает, базофилия протоплазмы постепенно ослабевает, специфическая зернистость увеличивается, причем у эозинофилов часто наряду с оксифильными зернами появляются отдельные базофиль-ные, неметахроматические зерна. Кроме того допускается, особенно при лейкозах, развитие миелоцитов непосредственно из индиферентной мезенхимной клетки (гемо-гистиобласт Феррата, рет.-энд. клетка) без промежуточного стадия гемоцитобласта. С другой стороны гемоцитобласт, подобно тому как у эмбриона, и во взрослом организме может развиваться из индиферентной мезенхимной клетки (см. ниже гетеротоп-ное кроветворение). Выход зрелых гранулоцитов в просвет синусоидов костного мозга объясняется их амебоидной подвижностью; здесь также может играть роль изменение давления крови в синусоидах, а следовательно и их просвета. Повышение интенсивности гранулопоэза наблюдается при многих инфекциях и особенно при миелозах. При нейтрофильном гиперлейкоцитозе усиливается выработка нейтрофилов и может наступить превращение жирового костного мозга в красный. В зависимости от силы и продолжительности вызывающей причины, клеточный состав костного мозга может резко меняться, и гетеропластиче-ский гранулопоэз значительно усиливается. Усиленное размножение эозинофильных миелоцитов и увеличенное образование их из гемоцитобластов наблюдаются при эози-нофильном гипер лейкоцитозе. При миелозах происходит превращение жирового костного мозга в миелоидный с развитием в паренхиме его более молодых гранулоцитов, а в острых случаях—гемоцитобластов. При хрон. инфекциях, интоксикациях и особенно при миелозах наряду с превращением жирового костного мозга в красный происходит развитие островков миелоидной ткани и в других местах организма (см. ниже гетеротопное кроветворение). Подавление гранулопоэза наблюдается при многих интоксикациях организма, при некоторых тяжелых инфекциях, при злокачественном малокровии. Костный мозг при этом беднеет гранулоцитами, и преобладающим элементом в нем может оказаться гемоцитобласт. Обычно это сопровождается дегенеративными изменениями гранулированных лейкоцитов и фокусами некроза паренхимы костного мозга. Отклонения от нормы в' развитии гранулоцитов могут состоять: 1) в недостаточной вы- работке специфической зернистости или даже полном отсутствии ее; 2) в неравномерном созревании ядра и протоплазмы; 3) в ненормальной дольчатости ядер гемоцитобластов (Ридеровские формы); 4) в развитии т. н. нейтрофильных близнецов, т. е. нейтрофилов, имеющих два ядра, совершенно-одинаково построенных. Особенно сильное отклонение от' нормы в гранулопоэзе наблюдается при острых миелозах: почти полное отсутствие гранулопоэза, причем преобладающим элементом в паренхиме костного мозга является гемоцитобласт. Иногда он имеет мелкие размеры, близко стоя по-своей морфологии к малому лимфоциту (ми-кромиелобласт).Из дегенеративных форм гранулоцитов заслуживают внимания дегенеративные формы палочкоядерных нейтрофилов, появляющихся в костном мозгу при так называемом дегенеративном ядерном сдвиге (Шиллинг). Образование лимфоцитов крови происходит в лимфоидной ткани, гл. обр. в лимф, узлах и селезенке, преимущественно-гомопластическим путем (рис. 7). Обычно считается, что малые лимфоциты крови происходят путем деления больших лимфоцитов, к-рые поэтому часто называются лимфобластами (Негели), реже—лим-фогониями Benda. Впрочем, по Максимову, гораздо чаще наблюдается размножение* средних—мезо-лимфоцитов. Деление лимфоцитов происходит в лимф, узлах, гл. образ.. в центре размножения фоликулов (рис. 7). Различают две фазы фоликулов лимфат. узла: деятельную и покойную. В течение первой—фоликул крупного размера содержит много больших и средних лимфоцитов' с многочисленными митозами; в течение второй—клеточных элементов в нем мало, преобладают малые лимфоциты. Г е т е р о-пластическое образование лимфоцитов происходит из индиферентных клеток мезенхимы. При этом фиксированная клетка мезенхимы делится митотически, давая происхождение круглым клеткам, имеющим светлые ядра и слабо базофильную протоплазму. При дальнейшем размножении последних получаются типичные средние лимфоциты. Этот процесс особенно выражен при переходе фоликула лимф, узла из фазы покоя в фазу деятельности (Максимов) . При развитии новых лимфатических фоликулов лимфоциты образуются из таких же индиферентных клеток мезенхимы. Это образование происходит преимущественно на месте бывших жировых долек. Генетические отношения фиксированных и свободных клеток лимфоидной ткани, по Максимову, можно схематически выразить следующим образом: ^ Средние и ->- Малые ^--' большие "*~ лимфо-Недиференцированный ' лимфоциты циты синцитий лимфоидной ~~-~^ ткани """-"-». Фиксиро- -> макро- ванные ги- фаги стиоциты При пат. условиях Максимов допускает кроме того возможность развития из больших и средних лимфоцитов элементов миелоидной ткани, из малых лимфоцитов—ми- кромиелоцитов, а из тех и других—моноцитов и макрофагов. Возможность происхождения моноцитов из свободных гистиоцитов (макрофагов) также допускается Максимовым. Патол. лимфопоэз, наблюдаемый при некоторых инфекциях, интоксикациях и особенно лимфаденозах, может состоять в довышенном образовании больших лимфоцитов—лимфобластов, у взрослого в крови нормально не встречающихся, далее—в ненормальной дольчатости ядер больших лимфоцитов (Ридеровские формы), в развитии лимфоцитов, почти лишенных протоплазмы или, наоборот, с обильным количеством последней, в отсутствии азурофильной зернистости, нормально встречающейся у одной трети лимфоцитов. Усиленный лимфопоэз сопровождается гиперплазией лимф, ткани с увеличением лимфатических узлов, селезенки, фоликулов слизистых оболочек, зобной .железы и с развитием новых лимфом в различных местах организма. Угнетение лим-•фопоэза происходит вследствие разрушения лимфойдной ткани лимфат. узлов; при этом кровь беднеет лимфоцитами (лимфо-гранулема, лимфосаркоматоз). Происхождение моноцитов крови не вполне выяснено. Имеется три главных взгляда: 1) моноциты происходят из рети-куло-эндотелиальн. аппарата путем мобилизации рет.-энд. клеток (Ашоф, Кийоно, Шиллинг); 2) моноциты развиваются в миелоид-ной системе из миелобластов (Негели); 3) моноциты образуются из лимфоцитов путем гипертрофии ядра и протоплазмы последних и представляют переходную форму от лимфоцита к макрофагу или полибласту (Максимов, Bloom). Такая гипертрофия лимфоцита происходит в кровеносных сосудах с замедленным током крови. Впрочем Максимов не отрицает происхождения моноцитов из ретикуло-эндотелия. Моноцитоз крови обычно сопровождается гиперплазией ретикуло-эндотелиального аппарата и мобилизацией ретикуло-эндотели-альных клеток, которые могут появляться в крови, особенно при некоторых хронических сепсисах. Эти клетки носят все признаки гистиоцитов или макрофагов и дают все возможные переходы к обычным моноцитам. Все это говорит за роль ретикуло-эндотелия в происхождении моноцитов; вместе с тем и лимфоцит, как показывают опыты эксплантации, может в самое короткое время перейти в моноцит. Тромбоцитопоэз (нормальный и патологический). У низших позвоночных и у птиц тромбоциты являются настоящими клетками, снабженными ядрами. У млекопитающих и человека кровяные пластинки представляют собой безъядерные образования, происходящие согласно исследованию Райта (Wright) из отделившихся частичек протоплазмы мегакариоцитов костного мозга. Тромбоцитопоэз происходит ловидимому таким образом, что мегакариоцит посылает в просвет синусоидов костного мозга амебоидные отростки азурофильной протоплазмы; от них отрываются мелкие частицы, к-рые и уносятся током крови в виде кровяных пластинок (рис. 1) (см. Виццоцеро бляшки). Впрочем нек-рые авторы до наст. времени придерживаются старых взглядов о происхождении кровяных пластинок, производя их то из протоплазмы лейкоцитов и эритроцитов то из ядер лейкоцитов.— Кровяная пластинка, по Шиллингу, представляет собой измененное ядро эритро-бласта, выталкиваемое из молодого эритроцита уже в кровяном русле. Повышенное развитие мегакариоцитов из гемоцитоблас-тов и усиленное функционирование их сопровождается повышением числа кровяных пластинок в крови и появлением гигантских форм (хлороз, миелозы). Иногда в кровь попадают части мегакариоцитов с обломками ядра. Некоторые патол. процессы сопровождаются тромбоцитопенией и появлением патологических пластинок с ненормальной зернистостью или отсутствием последней или с сильно выраженной базо-филией краевой зоны или пластинок ненормальных размеров (злокачественное малокровие, апластические анемии, «эссенциаль-ная тромбопения»). . Отношения между кроветворными органами и кровью. Морфолог, состав крови зависит от двух факторов: от производства форменных элементов и от их потребления, причем оба эти процесса находятся между собой в теснейшей взаимной связи и взаимно друг друга обусловливают. Повышение эритропоэза находит отражение в составе крови появлением регенеративных форм эритроцитов и эритробластов. Но при сильных и быстрых раздражениях костного мозга появление эритробластов в крови может быть вызвано простым вымыванием их из миелои-дной ткани вследствие нарушения кровообращения в ней. При мегалобластозе костного мозга в крови появляются мегалоци-ты и мегалобласты. Отсутствие в крови регенеративных форм эритроцитов при наличии тяжелой анемии позволяет предполагать отсутствие эритропоэза в костном мозгу (см. также Анемия). Отношения, существующие между лейкопоэзом и составом лейкоцитов крови, являются более сложными, так как распределение лейкоцитов в сосудистом ложе и процессы потребления лейкоцитов могут протекать независимо для каждого вида. При быстро развивающемся нейтрофиль-ном лейкоцитозе подвижные нейтрофиль-ные лейкоциты могут в самое короткое время эмигрировать из паренхимы костного мозга в кровеносные сосуды. При хрон. воспалительных процессах без осложнений, протекающих при почти полном отсутствии ядерного сдвига, костный мозг богат зрелыми и юными нейтрофилами, а при тяжелых острых инфекциях с сильным ядерным сдвигом он оказывается состоящим гл. обр. из промиелоцитов (Шиллинг). При лейкозах явления гиперплазии кроветворных органов в большинстве случаев находят отражение в лейкемич. составе крови, и по морфологии последней можно судить о морфологич. составе кроветворных органов, и наоборот. В.общем однако следует сказать, что состав крови далеко не всегда отражает процессы, протекающие в кроветворных органах; существует какой-то механизм, пока еще не выясненный, который, смотря по обстоятель- ствам, облегчает или затрудняет переход форменных элементов из костного мозга в периферическую кровь (костномозговой барьер). (См. также Лейкоцитозы, Лейкемия, Лейкоцитарная формула.) Гетеротопное К. При пат. условиях (ин-. фекции, интоксикации, тяжелые малокровия, миелозы) наблюдается возникновение миелоидной ткани в разных местах организма, т. н. экстрамедулярный миелопоэз. Миелоидная метаплазия как правило возникает сначала в селезенке, затем в печени, лимф, узлах, надпочечниках и других местах организма. При этом вначале появляются ми елоциты, за.теммегакариоциты, позднее и не всегда—эритробласты. Экстраме-дулярное происхождение миелоидных элементов в наст, время большинством объясняется соответствующей диференцировкой местных элементов (аутохтонная теория). Колонизационная теория, по к-рой миелоидная метаплазия возникает из клеток, заносимых током крови в различные места организма, в настоящее время большинством отвергается. Вопрос о том, из каких местных клеток развиваются элементы миелоидной ткани, является одним из наиболее спорных в гематологии. Часть авторов (Шридде, Герцог, Негели) производит их из клеток сосудистой стенки. Близко к этому стоит учение Феррата, Гоффа (Hoff), Сысоева и друг., приписывающее гемопоэ-тическую способность ретикуло-эндотелию. По Феррата, элементы миелоидной ткани развиваются из ретикуло-эндотелия, пройдя стадии гемогистиобласта и гемоцито-бласта. Меллендорф (Mollendorff) считает, что зернистые лейкоциты могут развиваться из фибробластов; это мнение впрочем встречает возражения большинства ученых. Доминичи, Доуни, Вайденрейх (Downey, Weidenreich), Максимов наделяют лимфоцитов гемопоэтическими способностями. По Максимову, гистиоциты и сосудистый эндотелий не обладают способностью к гемопоэзу, и экстрамедулярный миелопоэз происходит или за счет лимфоцитов крови и тканей или же из индиферентных мезенхимных клеток. Теории кроветворения. Вопрос о генетических отношениях форменных элементов крови друг к другу и клеточным формам соединительной ткани является и до наст. времени одним из самых спорных в гематологии. Здесь можно различать четыре главнейших направления, или четыре теории кроветворения: 1) унитарное направление. 2) умеренно-унитарное, 3) дуалистическое и 4) триалистическое. Унитарное учение (Гравиц, Максимов, Вайденрейх, Хлопин, Мясоедов) полагает, что все незернистые элементы крови и кроветворных органов, т. е. малые лимфоциты, большие лимфоциты, гемоцитобласты, а также (по нек-рым) и моноциты, являются недиференцированными клетками, способными при известных условиях давать происхождение зернистым лейкоцитам, эритроцитам и мегакариоцитам. Согласно этому учению гемоцитобласт (миелобласт) и большой лимфоцит способны давать происхождение одинаковым клеткам, а существую- I щие между ними биохим. и морфол. различия непостоянны и зависят исключительно от временных условий существования. Унитарное учение основывается главн. обр. на экспериментальных исследованиях, особенно—Максимова и его учеников. Согласно этому учению в раннем эмбриональном периоде все клетки мезенхимы обладают полнотой гемопоэтических способностей, во взрослом же организме эта способность сохраняется за отдельными недиференцированными клетками мезенхимы, к к-рым, по нек-рым, относится также и ретикуло-эндо-телий. По Максимову, часть клеток соединительной ткани односторонне диференциро-вана (фибробласты, эндотелий кровеносных и лимф, сосудов), часть (клетки ретикуло-эндотелиального аппарата и вообще гистиоциты) сохранила только некоторые способности (способность развиваться в фибробласты и в моноциты крови) и наконец часть сохранила все эмбриональные свойства с широкими гемопоэтическими возможностями (рисунок 2). Учение умеренных унитаристов (Паппенгейм) производит все элементы крови из общего родоначальника—лимфоидоцита (гемоцитобласта). Лимфоциты крови и лимфатической-ткани являются уже диферен-цированными элементами, неспособными к гемопоэзу. По учению дуалистов (Эрлих, Негели, Шридде) лимфоциты и зернистые лейкоциты имеют двух родоначальников—лимфобласта и миелобласта, к-рые ни при каких условиях не могут переходить друг в друга и различаются по морфологическим и биохимическим особенностям. Связующим звеном для элементов крови, по Негели, является индиферентная мезенхимная клетка, которая обладает всей полнотой гемопоэтических потенций. По триалистическому учению гистиоциты, как свободные, так и фиксированные, составляют 3-й независимый кроветворный орган, подобный миелоидной и лимф, системе организма. Этот орган производит моноциты крови, которые так. обр. по своему происхождению являются независимыми от лимфоцитов и зернистых лейкоцитов (Aschoff). Культура нормальной и пат. крови. Культура лейкоцитов крови in vitro, впервые произведенная Авроровым и Тимофеевским,. дала возможность выяснить нек-рые спорные вопросы гематологии. Зернистые лейкоциты нормальной крови как клетки с законченным развитием в течение нескольких дней подвергаются in vitro распаду, тогда как лимфоциты и моноциты развиваются в короткое время в фагоцитирующие поли-бласты или макрофаги. Этим подтверждается учение Максимова о роли незернистых лейкоцитов в происхождении клеточных: форм в воспаленной ткани. В более старых культурах макрофаги вытягиваются и постепенно переходят в типичные фиброциты, вырабатывающие тоно-фибрилы и колаге-новые волокна, — развивается настоящая соединительная ткань (Максимов). При своем развитии в полибласта лимфоцит крови I проходит через стадий моноцита, что гово- 54а рит за возможный генезис моноцитов из лимфоцитов (Максимов, Блюм, Тимофеев-ский и Беневоленская). Проявление гемо-поэтической способности со стороны лимфоцитов и моноцитов нормальной крови in vitro не происходит. В этом отношении эти клетки отличаются от гематоцитобласта (миелобласта) лейкемической крови: гемо-цитобласты в самое короткое время дифе-ренцируются in vitro в гранулоциты, реже—в эритробласты [см. отдельную таблицу (ст. 531—532), рисунки 3 и 4, и отдельную таблицу (ст. 659—660), рисунок 15] (Тимофеев-ский и Беневоленская). Сопоставление указанных двух факторов говорит не в пользу унитарного учения. Лит.—см. лит. к ст. Гематология и Костный мозг. А. Тдмофеевский.
Большая медицинская энциклопедия. 1970.
кроветворение — кроветворение … Орфографический словарь-справочник
КРОВЕТВОРЕНИЕ — гемопоэз (от гемо... и греч. poiesis изготовление, сотворение), размножение, развитие и созревание клеток крови в организме животных и человека в результате ряда по следоват. дифференцировок. Дифференцировка исходной полипотентной стволовой… … Биологический энциклопедический словарь
КРОВЕТВОРЕНИЕ — КРОВЕТВОРЕНИЕ, образование, развитие и созревание клеток крови. У взрослых особей млекопитающих и человека происходит в кроветворных органах костном мозге, селезенке, лимфатических узлах и вилочковой железе. Кроветворение непрерывный процесс,… … Современная энциклопедия
КРОВЕТВОРЕНИЕ — (гемопоэз) образование, развитие и созревание клеток крови. У беспозвоночных происходит в основном в полостных жидкостях и в самой крови или гемолимфе; у млекопитающих и человека в кроветворных органах. Кроветворение непрерывный процесс,… … Большой Энциклопедический словарь
кроветворение — гемопоэз Словарь русских синонимов. кроветворение сущ., кол во синонимов: 3 • гематопоезис (1) • … Словарь синонимов
Кроветворение — КРОВЕТВОРЕНИЕ, образование, развитие и созревание клеток крови. У взрослых особей млекопитающих и человека происходит в кроветворных органах костном мозге, селезенке, лимфатических узлах и вилочковой железе. Кроветворение непрерывный процесс,… … Иллюстрированный энциклопедический словарь
кроветворение — кроветворение. См. гемопоэз. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
кроветворение — гемопоэз Процесс размножения, развития и созревания клеток крови путем последовательных дифференциаций из исходной стволовой клетки; специализация клеток крови и снижение их способности к митозу при К. генетически детерминированы; у… … Справочник технического переводчика
кроветворение — я; ср. Процесс образования, развития и созревания клеток крови у животных и человека. ◁ Кроветворный, ая, ое. Относящийся к кроветворению. К ые органы. * * * кроветворение (гемопоэз), образование, развитие и созревание клеток крови.… … Энциклопедический словарь
Кроветворение — I Кроветворение (синоним гемопоэз) процесс, заключающийся а серии клеточных дифференцировок, в результате которых образуются зрелые клетки крови. Во взрослом организме существуют родоначальные кроветворные, или стволовые, клетки. Предполагают,… … Медицинская энциклопедия
КРОВЕТВОРЕНИЕ
18+
© Академик, 2000-2025
- Обратная связь: Техподдержка, Реклама на сайте
Экспорт словарей на сайты, сделанные на PHP, Joomla, Drupal, WordPress, MODx.